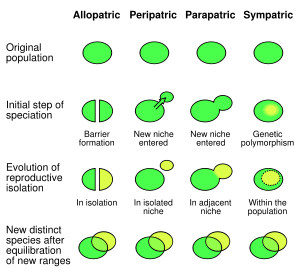
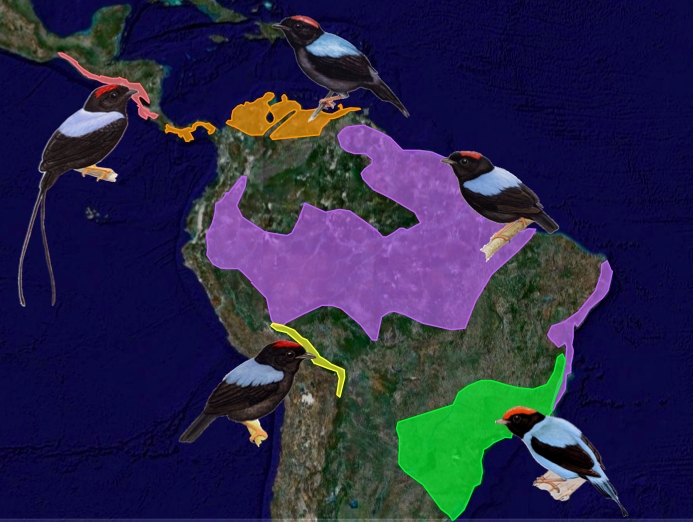
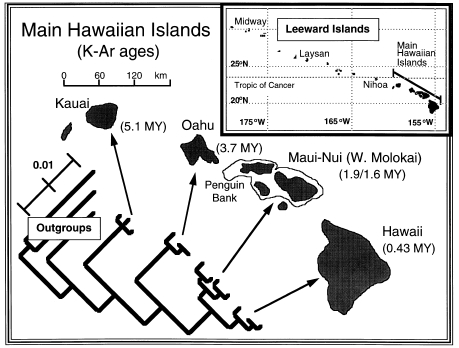
Some of the classics:
*Darwin, C. 1859. The Origin of Species by Means of Natural
Selection or the Preservation of Favoured Races in the Struggle for Life.
1962 edition by Collier Books, New York.
*Darwin, C. 1871. The Descent of Man, and Selection in
Relation to Sex. 1981 edition by Princeton University Press, Princeton,
NJ.
*Dobzhansky, T. 1937. Genetics and the Origin of Species.
Columbia University Press, NY.
Endler, J.A. 1977. Geographic Variation, Speciation and
Clines. Princeton University Press, Princeton, NJ.
*Fisher, R.A. 1958. The Genetical Theory of Natural Selection.
2nd edn. Dover Press, N.Y. (1st published in 1930 by Oxford University
Press).
*Gould, S.J. 1973. Ontogeny and Phylogeny. Harvard University
Press, Cambridge, Mass.
*Mayr, E. 1942. Systematics and the Origin of Species.
Columbia University Press, NY.
Otte, D. and J.A. Endler (eds.). 1989. Speciation and
its Consequences. Sinauer Associates, Sunderland, Mass.
Simpson, G.G. 1944. Tempo and Mode in Evolution. Columbia
Univ. Press, NY.
White, M.J.D. 1978. Modes of Speciation. W.H. Freeman,
San Francisco.
*Wright, S. 1978. Evolution and the Genetics of Populations,
Vol. 4: Variability Within and Among Natural Populations. University of
Chicago Press, Chicago.
Other references and literature cited:
Avise, J.C. 1994. Chapter 7: Speciation and hybridization.
In
Molecular Markers, Natural History and Evolution. Chapman and Hall, New
York.
*Avise, J. C., and K. Wollenberg. 1997. Phylogenetics
and the origin of species. PNAS 94: 7748-7755.
Ayala, F.J., and W.M. Fitch. 1997. Genetics and the origin
of species: An introduction. PNAS 94: 7691-7697.
Barton, N.H. 1996. Natural selection and random genetic
drift as causes of evolution on islands. Phil. Trans. Royal Soc. London
B 351: 785-795.
Barton, N.H., and G.M. Hewitt. 1989. Adaptation, speciation
and hybrid zones. Nature 341: 497-503.
Brumfield, R.T., and A.P. Capparella. 1996. Historical
diversification of birds in northwestern South America: a molecular perspective
on the role of vicariant events. Evol. 50: 1607-1624.
Brumfield, R.T., R.W. Jernigan, D.B. McDonald, and M.J.
Braun. 2001. Evolutionary implications of divergent clines in a manakin
(Manacus; Aves) hybrid zone. Evolution 55: 2070-2087.
*Bush, G.L. 1994. Sympatric speciation in animals � new
wine in old bottles. TREE 9: 285-288.
Clarke, B., M.S. Johnson, and J. Murray. Clines in the
genetic distance between two species of island land snails: how �molecular
leakage� can mislead us about speciation. Phil. Trans. Royal Soc. London
B 351: 773-784.
Coyne, J.A. 1994. Ernst Mayr and the origin of species.
Evol. 48: 19-30.
Coyne, J.A. 1995. Evolutionary biology: speciation in
monkeyflowers. Nature 376: 726-727.
Cracraft, J. 1983. Species concepts and speciation analysis.
Current Ornithology 1: 159-187.
Dall, S.R.X. 1997. Behavior and speciation. Trends Ecol.
Evol. 12: 209-210.
*Dawkins, R. 1976. The Selfish Gene. Oxford University
Press, N.Y.
*Dawkins, R. 1997. Human chauvinism: a review of "Full
House" by Stephen Jay Gould. Evolution 51: 1015-10120.
Endler, J.A., and M. Théry. 1996. Interacting effects
of lek placement, display behavior, ambient light, and color patterns in
three Neotropical forest-dwelling birds. Am. Nat. 148: 421-452.
*Feder, J.L., C.A. Chilcote, and G.L. Bush. 1988. Genetic
differentiation between sympatric host races of the apple maggot fly Rhagoletis
pomonella. Nature 336: 61-64.
*Fleischer, R.C., C.E. McIntosh, and C.L. Tarr. 1998.
Evolution on a volcanic conveyor belt: using phylogeographic reconstructions
and K-Ar-based ages of the Hawaiian Islands to estimate molecular rates.
Mol. Ecol. 7: 533-545.
*Gallardo, M. H., J. W. Bickham, R. L. Honeycutt, R. A.
Ojeda, and N. Kohler. 1999. Discovery of tetraploidy in a mammal. Nature
401: 341.
Gascon, C, J.R. Malcolm, J.L Patton, M.N. da-Silva, J.P. Bogart, S.C. Lougheed, C.A. Peres, S. Neckel,
and P.T. Boag. 2000. Riverine barriers and the geographic distribution of Amazonian species.
Proc. Natl. Acad. Sci. USA 97: 13672-13677.
*Gould, S.J. 1997. Self-help for a hedgehog stuck on a
molehill: a review of "Climbing Mount Improbable" by Richard Dawkins. Evolution
51: 1020-1024.
*Grant, P.R. 1986. Ecology and Evolution of Darwin�s Finches.
Princeton University Press, Princeton, NJ.
*Grant, P.R., and B.R. Grant. 1996. Speciation and hybridization
in island birds. Phil. Trans. Royal Soc. London B 351: 765-772.
Grant, P.R., and B.R. Grant. 1997. Genetics and the origin
of bird species. PNAS 94: 7768-7775.
Green, D.M., T.F. Sharbel, J. Kearsley, and H. Kaiser.
1996. Postglacial range fluctuation, genetic subdivision and speciation
in the western North American spotted frog complex, Rana pretiosa.
Evol. 50: 374-390.
Haldane, J.B.S. 1966. The Causes of Evolution. Cornell
University Press, Ithaca, New York.
Hostert, E.E. 1997. Reinforcement: a new perspective on
an old controversy. Evol. 51: 697-702.
Johnson, N.K.1995. Speciation in vireos. I. Macrogeographic
patterns of allozymic variation in the Vireo solitarius complex
in the contiguous United States. Condor 97: 903-919.
Koeslag, J.H. 1995. On the engine of speciation. J. Theor.
Biol. 177: 401-409. �
Krutovskii, K.V., and F. Bergmann. 1995. Introgressive
hybridization and phylogenetic relationships between Norway, Picea abies
(L) karst, and Siberian, P. obovata ledeb, spruce
species studied by isozyme loci. Heredity 74: 464-480
*Lande, R. 1981. Models of speciation by sexual selection
on polygenic traits. PNAS USA 78: 3721-3725.
Liou, L.W., and T.D. Price. 1994. Speciation by reinforcement
of premating isolation. Evolution 48: 1451-1459.
Lovette, I.J., E. Bermingham, and R.E. Ricklefs. 2001. Clade-specific morphological diversification and adaptive radiation in
Hawaiian songbirds. Proceedings of the Royal Society of London: Biological Sciences 269:37-42.
*Meyer, A., L.L. Knowles, and E. Verheyen. 1996. Widespread
geographical distribution of mitochondrial haplotypes in rock-dwelling
cichlid fishes from Lake Tanganyika. Molecular Ecology 5(3): 341-350.
Mooers, A.O., and A.P. Møller. 1996. Colonial breeding
and speciation in birds. Evol. Ecol. 10: 375-385.
Orr, H.A., and L.H. Orr. 1996. Waiting for speciation:
the effect of population subdivision on the time to speciation. Evol. 50:
1742-1749.
Orr, H.A. 1997. Haldane's Rule. Ann. Rev. Ecol. Syst.
28: 195-218.
*Orr, M.R., and T.B. Smith. Ecology and speciation. Trends
Ecol. Evol. 13: 502-506.
Parsons, T.J., S.L. Olson, and M.J. Braun. 1993. Unidirectional
spread of secondary sexual plumage traits across an avian hybrid zone.
Science 260: 1643-1646.
Pearson, S.F., and S. Rohwer. 2000. Asymmetries in male
aggression across an avian hybrid zone. Behav. Ecol. 11: 93-101.
Rice, W.R., and E.E. Hostert. 1993. Laboratory experiments
on speciation: What have we learned in forty years? Evol. 47: 1637-1653.
Rieseberg, L. H., A. Widmer, A.M. Arntz, and J.M. Burke.
2002. Directional selection is the primary cause of phenotypic diversification.
PNAS USA 99: 12242-12245.
*Roderick, G.K, and R.G. Gillespie. 1998. Speciation and
phylogeography of Hawaiian terrestrial arthropods. Molecular Ecology 7:
519-531.
Rohwer, S., and C. Wood. 1998. Three hybrid zones between
Hermit and Townsend's warblers in Washington and Oregon. Auk 115: 284-310.
Savolainen, V., M.-C. Anstett, C. Lexer, J. Hutton, J.C. Clarkson, M.V. Noroup, M.P. Powell, D. Springate, N. Salamin, and W.J. Baker.
2006. Sympatric speciation in palms on an oceanic island. Nature 441: 210-213.
Schliewen UK, Tautz D, Paabo S. 1994. Sympatric speciation
suggested by monophyly of crater lake cichlids. Nature 368: 629-632.
*Schluter, D. 2001. Ecology and the origin of species. Trends in Ecology and Evolution 16:372-380.
*Schluter, D., and T. Price. 1993. Honesty, perception
and population divergence in sexually selected traits. Proc. Roy. Soc.
London B 253: 117-122.
*Slatkin, M. 1996. In defense of founder-flush theories
of speciation. Am. Nat. 147: 493-505.
*Smith, T.B., R.K. Wayne, D.J. Girman, and M.W. Bruford.
1997. A role for ecotones in generating rainforest biodiversity. Science
276: 1855-1857.
Sorenson, M.D., K.M. Sefc, and R.B. Payne. 2003. Speciation by host switch in parasitic indigobirds. Nature 424: 928-931.
Stein, A.C., and J.A.C. Uy. 2006. Unidirectional introgression of a sexually selected trait across an avian hybrid zone: a role for female choice? Evol. 60: 1476-1485.
*Tegelström, H., and H.P. Gelter. 1990. Haldane�s
rule and sex biased gene flow between two hybridizing flycatcher species
(Ficedula albicollis and F. hypoleuca, Aves: Muscicapidae).
Evol. 44: 2012-2021.
Templeton, A.R. 1980. Modes of speciation and inferences
based on genetic distances. Evol. 34: 719-729.
*Templeton, A.R. 1996. Experimental evidence for the genetic-transilience
model of speciation. Evol. 50: 909-915.
True, J.R. et al. 1997. Quantitative genetic analysis
of divergence in male secondary sexual traits between Drosophila simulans
and Drosophila mauritania. Evol. 51: 816-832
Turelli M. 1998. The causes of Haldane's rule. Science
282, 889-891.
*Turner, G.F., and M.T. Burrows. 1995. A model for sympatric
speciation by sexual selection. Proc. Roy. Soc. London B 260: 287-292.
Turner, J.R.G., and J.L.B. Mallet. 1996. Did forest islands
drive the diversity of warningly coloured butterflies? Biotic drift and
the shifting balance. Phil. Trans. Royal Soc. London B 351: 835-845.
*Verheyen, E., L. Rüber, J. Snoeks, and A. Meyer.
1996. Mitochondrial phylogeography of rock-dwelling cichlid fishes reveals
evolutionary influence of historical lake level fluctuations of Lake Tanganyika,
Africa. Phil. Trans. Royal Soc. London B 351: 797-805.
*Wade, M.J., and C.J. Goodnight. The theories of Fisher
and Wright in the context of metapopulations: when nature does many small
experiments. Evol. 52: 1537-1553.
*West Eberhard, M.J. 1983. Sexual selection, social competition
and speciation. Quart. Rev. Biol. 58: 155-183.
*Zeh, D.W., and J.A. Zeh. 1994. When morphology misleads:
interpopulation uniformity in sexual selection masks genetic divergence
in harlequin beetle-riding pseudoscorpion populations. Evolution 48: 1168-1182.
Zink, R.M. 1994. The geography of mitochondrial DNA variation,
population structure, hybridization, and species limits in the fox sparrow
(Passerella iliaca). Evol. 48: 96-111.
*Zink, R.M., and M.C. McKitrick. 1995. The debate over
species concepts and its implications for ornithology. Auk 112: 701-719.
Return to top of page