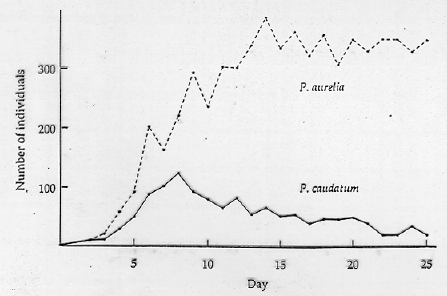
Fig. 19.1. Competitive exclusion of one species of the ciliate protozoan Paramecium by another.
Data from Gause (Gause, G.F. 1934. The Struggle for Existence. Williams and Wilkins, Baltimore, MD.).
Lecture 19 (Mon. 4-Mar-13) Population dynamics involving two species -- Interspecific competition.
Required reading: Gotelli Chapter 5 (see WyoWeb)
Return to Main Index page Go back to lecture 18 27-Feb-13 Go forward to lecture 20 Wed. 6-Mar-13
Thus far in the course we have dealt with the dynamics of single populations as if they were acting in isolation -- with dynamics that depended only on their own internal dynamics. We looked at various forms of population growth, the effects of age structure and sex ratio and the nature of intraspecific competition. For most of the remainder of the course we will focus on how populations interact in multi-species interactions -- we will look at interspecific competition, predation, harvesting and host-pathogen and disease dynamics. Let's begin with a chart that shows the range of kinds of interactions.
Classifying interspecies interactions
Table 19.1. Classification of the major sorts of interspecific interactions (omitting 0/0).
We will focus almost exclusively on predation, host/parasite interactions, and competition.
Description Effect on Species 1 Effect on Species 2 Example Mutualism + + Lichens (fungus/alga); cleaner/reef fish; ants/acacias Commensalism + 0 Epiphyte/cloud forest tree *Predation + - Predator/prey; host/parasite Amensalism 0 - Allelopathy (poison competitors) *Competition - - Interspecific competition; niche overlap/ resource overlap
* categories that will be major topics for the purposes of this course.
We will begin by examining some of the dynamics of interspecific competition.
Remember back, also, to a way of classifying competition by the mechanism -- exploitation vs. interference competition. Exploitation competition involves using up a resource (so that it is no longer available to another individual or species). Interference competition involves behavioral interactions that keep others from gaining access. For some sedentary species a third term is useful -- preemptive competition occurs when individuals use space that then becomes unavailable to others (barnacles in the rocky intertidal, plants; I suppose you could think of cavity occupancy as preemptive if you like). Most of the competition we will address will involve exploitation -- individuals or species competing for a limited and depletable resource.
Niches and resource partitioning: One of our major questions will be to look at where and when competitors can coexist. To understand that we need to begin with a review of niche theory and the idea of competitive exclusion.
From introductory ecology you should remember the idea of a niche. One way to think of it is as the ways that an organism uses resources. This is an especially appropriate way to think of it in the context of competition, because the main feature of competition is limited resources for which animals or plants need to compete. Let's run through a few examples to get the idea.
The niche of several species of desert rodents as well as ants might entail storage and consumption of seedsKinds of resources: food, shelter, water, hosts (for parasites), nesting/denning sites.
The niche of several species of woodpeckers, owls, chickadees and other birds, as well as flying squirrels, might entail using tree cavities for nesting or roosting.
The niche of ungulates might include the types and times of use of browse and forage.
The competitive exclusion principle says that if two species have almost completely overlapping niches they cannot continue to coexist. One of the two species will outcompete the other and persist. The other will go locally extinct. The classic experiments were by Gause using two species of Paramecium, a ciliate protozoan.
Even though both species initially grew exponentially, the outcome was always that one species came to dominate the culture and eventually drove the other to extinction.
Fig. 19.1. Competitive exclusion of one species of the ciliate protozoan Paramecium by another.
Data from Gause (Gause, G.F. 1934. The Struggle for Existence. Williams and Wilkins, Baltimore, MD.).
That is not, however, the most interesting possible result. The most interesting result is when the two species coexist at intermediate densities. That is, they compete, they have an effect on each other, but neither can completely dominate. In that case, we have coexistence. Another interesting case is when only one species persists, but the persistence depends on initial density. That is, rather than "Species 1 always wins", Species 1 wins only if it starts out at a higher density than Species 2. We will explore those kinds of interactions shortly by using paired sets of logistic equations.
Often, we need to consider several different kinds of niche elements simultaneously -- the idea of a multidimensional niche. For example, hawks (diurnal) and owls (nocturnal) have somewhat similar niches in terms of prey, but divide up the resources by foraging at different times (day vs. night) and by tending to use different sensory modalities (vision vs. hearing). The way that resources are divided up among sets of species with somewhat different but sometimes overlapping needs is called resource partitioning.
Fundamental vs. realized niche -- in the absence of competitors (that absence may not actually occur in the real world) a species will tend to have a characteristic niche width. This is called its fundamental niche. In the presence of competitors, however, it may be restricted to a narrower portion of its fundamental niche -- that is its realized niche.
Limiting similarity -- if two potential competitors start far apart in their use of resources, it means that some resources won't be used. If they overlap too much, competition may hurt members of both species -- (complete overlap should lead to the competitive exclusion described earlier). A balance or tension results -- between too little overlap (and therefore inefficient use of the available resources) on the one hand, and too much overlap (and therefore intense competition possibly even leading to competitive exclusion) on the other hand. The problem of limiting similarity is "how different do the niches of two species have to be in order for them to coexist?" The tension between greater similarity (driven by the pressure for efficiency) and less similarity (driven by competition) may largely determine the observed degree of similarity among competitors.
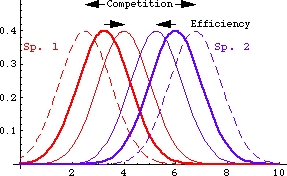
Example of resource partitioning adjustments: interplay between avoiding competition and using resources efficiently. Consider first a two-dimensional plot (Fig. 19.2, below) along an axis describing a niche dimension -- it is probably easiest to think of the X-axis as being related to use of a food or other resource (say seed size for desert rodents for example). The Y-axis is some measure of response (growth, fitness) to that resource distribution (or the proportion of animals using that portion of the resource spectrum).
The outcome of interspecific competition on an evolutionary scale can have two interesting outcomes -- character displacement and competitive release.
Resource dimension (e.g., seed volume in mm3).Fig. 19.2. Niche overlap diagram for two species using similar resources (e.g., seeds of desert plants). The X-axis depicts some measurable aspect of a niche dimension, such as food resources (e.g., seed size). The Y-axis shows the frequency of use. In the absence of its competitor, each species uses a range of resources that represents its fundamental niche (thickest solid curves). Competition will tend to select for a reduction in overlap (outward pointing arrows, dashed resource use curves that are further from each other). Selection for efficient use of resources (arrows pointing inward) will favor even greater overlap (thin, solid curves that are closer to each other). Note that each species has a peak mean value with a (bell-shaped, normal) distribution of use of larger and smaller seeds.
Character displacement. Given some degree of overlap in resource use, species should be under selection (and should respond to that selection) by evolving niche dimensions with lower overlap. It has been difficult to demonstrate character displacement in nature but a few reasonable examples exist.
Character release and character displacement are flip sides of the same coin. If the starting state is two species together, and then one or both occur without the competitor, then in that unusual "free-of-competition" location we would talk of character release (species niche width likely to expand to a broader fundamental niche). If, however, the species usually occur in different locations, then in the unusual location where they co-occur we might see a shift away from each other, and we would term that character displacement (and the realized niche width will tend to narrow and perhaps shift its mean away from the fundamental niche).
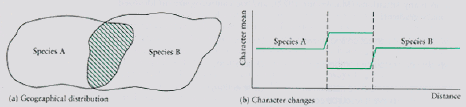
Fig. 19.3. Diagrammatic representations of the process of character displacement. a) Two species have overlapping geographic ranges (the overlap is shown by green cross-hatching). b) In the area of overlap, the two species are more different (in some character such as bill depth or wing length) than they are where they occur in the absence of the competitor. This mutual shift of a character in the presence of a competitor is called character displacement. In this case Species A has developed a larger character, whereas Species B has shifted to a smaller character. [From Krebs Ecology, 4th edn.] Character release. If a species has had a competitor that then disappears it is likely to broaden its niche width to use the resources that were previously used by the competitor. One of the best places to see this is in the Galapagos Islands where several species of Darwin's finches (genus Geospiza) sometimes co-occur on one island but do not co-occur on other islands. In that case, the tendency will be for the species without the competitor to use a broader range of resources than when it occurs with the competitor. [Note that character displacement and competitive release are flip sides of the same coin -- the former results when two competitors come together, the latter when formerly co-existing competitors no longer co-occur. You have to know "which came first" in order to say whether the transition is displacement or release].
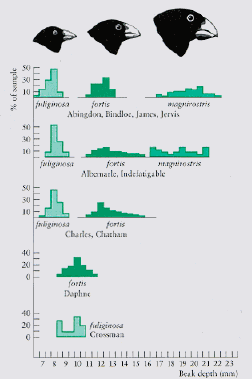
Fig. 19.4. Character release (and/or character displacement) in differing combinations of species of Darwin's finches (genus Geospiza). On the X-axis are beak depth classes for three species of finches. On the Y-axis are the numbers of finches with the given beak depths for a series of islands in the Galapagos. Note, for example, that Geospiza fuliginosa finches on Crossman Is. occur alone. There they have a larger range of beak sizes and that the mean is larger, than on the top three sets of islands, where they co-exist with two other competitors. The key point in this graph is the contrast between the three islands where fuliginosa and fortis co-occur and the two islands (bottom) where each occurs by itself. The lower cases suggest wider, and shifted, and highly overlapping fundamental niches for fortis and fuliginosa. (From: Krebs. Ecology, 4th Edn.)
The rest of this "lecture" will be web only.
In a classic paper Schoener (1974) looked at a wide range of published studies of interspecific competition and came up with some generalizations about how interspecific competition works:
1) Habitat more important than food-type, which is more important in turn than temporal separation as a way of avoiding intense competition by using a different niche componentReferences:
Example: Warblers using different parts of the tree (structural separation)2) Predators separate more often by being active at different times of the day than do other kinds of animals.
Examples: Hawks and owls, lions and leopards3) Terrestrial poikilotherms(= ectotherms, e.g., lizards, snakes) relatively often partition food by being active at different times of the day.
Example: Lizards (some active in cool part of day, others at hottest times)4) Vertebrates segregate less by seasonal activity than do lower animals
Example: Mayflies and other stream invertebrates will often emerge at different times of the year from the emergence times of their potential competitors5) Segregation by food type or foraging strategy is more important for animals feeding on food that is large in relation to their own size than it is for animals feeding on small food items.
Example: Large cats with antelope types (sit-and-wait vs. active chase strategies) vs. shorebirds with invertebrate prey (where many species will use the same prey items, such as polychaetes).
Schoener, T.W. 1974. Resource partitioning in ecological communities. Science 185: 27-39.§§§§§§§§§§§§§§§ §§§§§§§§§§§§§ §§§§§§§§§§§§§§ §§§§§§§§§§§§§ §§§§§§§§
Return to top of page Go forward to lecture 20 Wed. 6-Mar-13