
Grasshoppers of Wyoming and the West
Entomology
Northern Grasshopper
Melanoplus borealis (Fieber)
 |
Distribution and Habitat
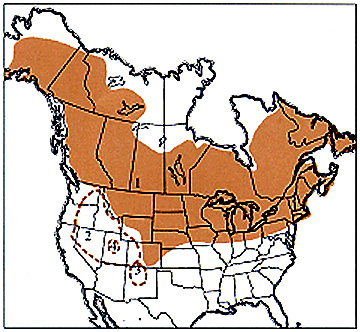 M. borealis continental distribution map >
M. borealis continental distribution map >The northern grasshopper, Melanoplus borealis borealis, ranges widely in the north of North America. The species inhabits both lowland and mountain sites, living in a variety of habitats that range from arctic tundra, wet bogs, and swamps to moist mountain meadows. In the latter, large populations consisting almost entirely of the northern grasshopper irrupt sporadically.
Economic Importance
Because the northern grasshopper feeds preferentially on certain forbs and variably on certain grasses, a clear picture of its economic importance on grazing lands eludes precise description. In mountain meadows of Montana and Wyoming this grasshopper increases sporadically to densities as high as 60 young adults per square yard. It thus has the potential of damaging range forage; indeed, experienced entomologists have reported serious damage. In 1955 in Utah's Beaver Mountain, Fishlake National Forest, grasshopper survey entomologists reported a population of the subspecies M. borealis palaceus, estimated at 50 young adults per square yard, that destroyed reseeded meadow. High densities of this grasshopper have also been reported in the mountains of Idaho and Oregon. A small outbreak of the northern grasshopper in Wyoming occurred in 1995 in the Big Horn Mountains. One infested meadow of approximately 100 acres supported a population estimated at 20 young adults per square yard. A visual estimate of the vegetation indicated that 90 percent had been grazed by August 9. A method of sorting out the proportions consumed by each of the two chief herbivores, cattle and grasshoppers, appeared impossible. Observations of damage in nature are usually confounded by the presence of more than one herbivore. A study with adequate controls is needed to determine the exact impact under various circumstances. The problem is complex as the northern grasshopper is also beneficial when it feeds on its preferred host plants of lupine and loco. Grasshopper management specialists in Wyoming have postulated that in forb-rich meadows, damage to range grasses may be much less than might be expected from high densities of this species.
The northern grasshopper is a medium-sized species. Males collected from a meadow in the Big Horn Mountains of northern Wyoming averaged 231 mg live weight and females 392 mg (dry weight: males 68 mg, females 111 mg). Live weight of Alaskan males one to two days after emergence averaged 254 mg and females 377 mg.
Food Habits
Northern grasshoppers feed principally on forbs but they may also feed at times on certain grasses. In the crops of adults collected in the mountain meadows of Wyoming, high percentages of lupine and loco have been found (Table 1). Other forbs selected include dandelion, Cerastium arvense, species of thistle (Cirsium), and cinquefoil (Potentilla). As do many species of grasshoppers, the northern grasshopper will feed on arthropods, probably dead or dying grasshoppers, when they have the opportunity. A bluegrass, Poa sp, has been found in large amounts in crops of the northern grasshopper collected in one of three Big Horn Mountain sites (Table 1).
Near Fairbanks, Alaska, in an open field, the northern grasshopper has been observed feeding on four forbs: dandelion, red clover, Parnassia palustris, and Petasites frigidus. In laboratory tests these species were selected for food, confirming their status as preferred host plants in the Alaskan habitat.
In 1962 in the sand prairie of southeastern North Dakota, flood waters trapped a population of northern grasshoppers with a large number left in the center of an ongoing study area. Analysis of crop contents revealed that the grasshoppers were feeding on seven forbs, five grasses, and one sedge. Favored foods in the order of percent ingested were Kentucky bluegrass, 53; western ragweed, 21; Missouri goldenrod, 15; and leadplant, 9.
In two-choice food preference tests of adults collected from a heavily infested meadow in the Big Horn Mountains of Wyoming, the grasshoppers preferred dandelion to white clover, alfalfa, silky loco, young wheat plants, downy brome, Kentucky bluegrass, western wheatgrass, and Idaho fescue. Of particular interest is the result of pairing dandelion and western wheatgrass. After six hours, 75 percent of the dandelion was consumed but less than 1 percent of the wheatgrass. After 22 hours all of the dandelion had been consumed and 20 percent of the western wheatgrass. Evidently the grasshoppers will feed on wheatgrass when other food is lacking. After six hours 98 percent of dandelion and none of Idaho fescue were consumed. In contrast, Kentucky bluegrass and downy brome were consumed at the same time as dandelion but in unequal amounts. At the end of four hours, 90 percent of dandelion and 25 percent of Kentucky bluegrass were consumed; at the end of six hours 20 percent of dandelion and 10 percent of downy brome.
In its mountain meadow habitat a few observations have been made of the northern grasshopper’s method of attacking its food plant. A male was seen to rise from a horizontal position on the ground onto the lower green leaves of field chickweed, Cerastium arvense. Resting diagonally on the plant with hindlegs on the ground supporting the body, the male consumed a half-inch long leaf from tip to base. Another observation was of a female attacking a forb sprout. Oriented horizontally on the soil surface and over the plant, the female ate in two minutes the entire sprout from tip to below ground level.
No observation of the grasshoppers feeding on lupine (Lupinus wyethi) was made, but damage to leaves showed feeding from tip to base and gouging from edges. Evidently the grasshoppers either climbed 3 to 10 inches on the stems or they jumped onto the plant to reach the leaves.
Dispersal and Migration
The northern grasshopper possesses long wings, enabling it to fly and disperse. Observations of the species in a 4 acre meadow near Fairbanks, Alaska, provide insights into its dispersal behavior. Nymphs were concentrated in loose groups around areas where the eggs had hatched, but upon the acquisition of wings, the grasshoppers dispersed progressively into wider areas. When temperatures rose from 74° F to 95° F, the adults flew frequently and covered wide areas in relatively short periods. Another example of this grasshopper's powers of dispersal happened fortuitously when one-half of the meadow (2 acres) was mowed in mid July, 1967. Five days later the density of adults in the unmowed half did not differ greatly from that in the mowed. Ten days later, however, the density in the mowed half more than doubled. Finally, 15 days after mowing the population was concentrated in the mowed section. Because of the very short vegetation in this section the ground temperature was 4 to 6° F higher than in the natural habitat. Presumably, the grasshoppers moved because they preferred the higher temperatures.
A record of dispersal or of migratory flight was the discovery of a male northern grasshopper on the ice of Grasshopper Glacier in the Crazy Mountains of Montana, August 1988. The wings of the male were long, surpassing the apex of the hind femur by 5 mm. Individuals of the species were present in the adjacent meadow, so it is unknown whether the male came from close by or from a distant meadow.
In meadows of the Big Horn Mountains, flushed flight by the northern grasshopper is silent, short (1 1/2 to 3 feet), and at heights of 2 to 4 inches. The flight starts usually from the ground and ends on the ground, with the grasshopper facing away from the intruder. In the meadow at Fairbanks, Alaska flushed grasshoppers flew a distance of 1.6 to 5 feet at heights of 1 to 2 feet. On very warm, dry, and windless days, some adults flew 3 feet high and covered a distance of 10 to 20 feet.
Identification
The northern grasshopper is a dark insect of medium size possessing wings of variable length. The wings of males usually range from slightly short of the apex of the hind femur to slightly beyond; the wings of females are usually shorter covering three quarters of the abdomen (Fig. 6 and 7). The shape of the male cercus is diagnostic of the species (Fig. 9). It is short, wide at the base, gently curved upward, and narrows toward a blunt apex. The arms of the furcula are long, reaching about halfway on the supraanal plate. The subgenital plate curves dorsally, terminating in a blunt truncate end. The medial area of the hind femur is usually entirely black, the upper marginal area is usually fuscous, the lower marginal area is red but sometimes yellow, and the hind tibia is red (Fig. 8). The abdominal venter is usually pale gray or pale green, but occasionally yellow or two-toned, each sternum being fuscous in the anterior half and olive in the posterior.
The nymphs are identifiable by their color patterns, shape, and structures (Fig. 1-5). The first instars are distinctively black except for a narrow mid dorsal brown band and a light crescent on the side of the head and pronotum. Figure 1 pictures an unusually light first instar.
1. Head with face nearly vertical and colored black, brown, tan, or green; yellow or ivory crescent on side of head and lateral lobe of pronotum; compound eye with many light spots, lower half darker than upper; narrow light line and broad black bar behind compound eye.
2. Pronotum brown or green and usually with dark brown or fuscous spots. Medial area of hind femur all black in instars I an II, broad dorsal stripe in instars III to V; inner medial area in instars I and II all black and occasionally marked by a light bar apically; in instar III like instar II or the black area may be interrupted by two light bars, an apical and a middle one; in instarts IV and V the inner black area becomes a dark dorsal ensiform stripe (elongate triangular with base toward apex of hind femur), light ventrally. Hind tibia is black in instar I with a light annulus proximally, in instar II hind tibia is black or fuscous and tan. In instars III to V hind tibia is usually dark tan with front fuscous.
3. General body color of instars II to V brown less often green, venter generally olive or each sternum of the abdomen with front half fuscous and rear half olive.
Hatching
In the grasshopper assemblages of mountain meadows, the northern grasshoper hatches relatively early along with several other high-altitude species, including Aeropedellus clavatus, Bruneria brunnea, Camnula pellucida, Melanoplus alpinus, and M. bruneri. Before hatching in spring, the eggs of the northern grasshopper require two winters in the soil to break diapause. The species is one of several that have a two-year life cycle. In the early 1950s field entomologists postulated that this grasshopper required more than one year to complete its life cycle when they observed reinfestations of areas a year after treatment and were unable to explain the results by poor kills or migration of adults. Subsequently, entomologists conducted field and laboratory experiments that showed the eggs required two years in the soil before hatching.
A study of the life cycle of the northern grasshopper in the Big Horn Mountains of Wyoming in 1994 and 1995 revealed that eggs begin to hatch from June 1 to July 1. The exact date depended on time of snow melt, warming of the soil, and elevation of site. On 4 May 1994, Powder River Pass (elevation 9,660) was 85 percent covered with snow. By May 24 the site became clear of snow, and exactly one month later the eggs of the northern grasshopper began to hatch. A late spring in 1995 delayed the start of hatch at this site until July 1. In this year hatching continued for two weeks.
In a meadow site near Fairbanks, Alaska (elev. 450 feet) eggs of the northern grasshopper began hatching June 10 in both 1967 and 1968 and continued for 10 to 15 days. The eggs did not hatch unless they were submerged in melt water in spring.
Nymphal Development
The nymphs grow rapidly, completing their development to the adult stage in about one month. Calculated from first hatch to first adults, nymphs in the Bald Ridge site (elevation 9,185) of the Big Horn Mountains, completed development in 29 days; while in the Powder River Pass site (elevation 9,660 feet) they completed development in 31 days. In the meadow at Fairbanks, Alaska, nymphs took 30 to 32 days to become adults.
Adults and Reproduction
In meadows of the Big Horn mountains, adults may appear as early as the first week of July or as late as the first week of August. They usually remain in the habitat in which they developed as nymphs and some survive into mid September. Maturation in nature has not been studied but rearing of the species in a laboratory at Fairbanks, Alaska has provided information on this important feature of the adult life history. The adults emerged between July 10 and August 3; the first mating was observed on July 28 but mating did not become common until August 7. Sexual maturity was attained about three weeks after molting to adulthood. In laboratory cages mating usually occurred during the warmest part of the day and pairs remained in copulo for 10 to 15 minutes. Oviposition began 8 to 10 days after mating and lasted from mid August until October 1.
No record of females ovipositing in their natural habitat has been made. Under laboratory conditions females readily oviposit into bare soil. The pods, 3/4 to 7/8 inch long and slightly curved, contain 12 to 16 eggs (Fig. 10). The egg section is 3/8 inch long and lies in the soil at a depth equal to the length of the pod. Eggs are two-toned tan and yellow and 4 to 4.8 mm long.
Population Ecology
The northern grasshopper has a history of irrupting in meadows of the Rocky Mountains. Survey records from the Big Horn Mountains of Wyoming have revealed populations as dense as 60 young adults per square yard. The species enjoys a high frequency of occurrence in the many disjunct meadows of the Rocky Mountains. For example, of 19 meadow sites investigated in 1994 and 1995, nine harbored individuals of the northern grasshopper and ten were free of the species (47 percent frequency). Densities of three sites were estimated to be 20, 8, and 3 young adults per square yard while densities of the other six occupied sites were all less than 0.1 per square yard. In outbreak years the species appears to have a higher frequency of occurrence. In 1963, an outbreak year, all of 14 sites surveyed were infested and dominated by the northern grasshopper. The density of assemblages in these sites averaged 49 young adult grasshoppers per square yard. We may infer that the northern grasshopper comprised over half of these densities and it therefore comprised an average of 25 or more young adults per square yard. In 1964 the numbers of northern grasshopper were sparse and their dominance was relinquished to Camnula pellucida and Melanoplus sanguinipes.
Because only limited studies of the population ecology of the northern grasshopper have been conducted and none for more than three years, we do not know how many years it takes for populations to increase to outbreak numbers, nor how long outbreak populations persist after they appear. We also do not know how long the intervals of low numbers last. Survey data indicate that an infested site may have high densities in successive years or due to the biennial life cycle of the species particular sites may have high numbers one year and low or none the next. Evidently a multiplicity of physical factors are affecting the growth, survival, and decline of populations of the northern grasshopper.
Daily Activity
The northern grasshopper dwells chiefly on the ground in meadow habitats with adequate bare soil for basking. Its nightly refuge has yet to be determined with certainty. An observation made 10 August 1995 at 7:30 a.m. DST in a meadow of the Big Horn Mountains (sun hid by clouds, 40° F ground surface, 50° F air) revealed a female sitting quietly on and parallel to a dry stem of grass that leaned at a 10° angle to the ground. The female rested at a height of 2 inches above ground and was sheltered in a thick midgrass canopy. On clear days grasshoppers leave shelters early in the morning and move to sunny spots on bare soil or litter to bask. Observations made in August 1995 from 9 to 11 a.m. DST show that the basking grasshoppers orient themselves horizontally on the soil surface, turn a side perpendicular to the sun, and lower the associated hindleg to expose the abdomen fully to the sun’s rays. On clear days basking ends between 10 and 11 a.m. when ground temperatures have increased to 85° F and air temperatures exceed 52° F. The grasshoppers may turn on the ground and face directly away or into the sun and rest, or they may become active--walking, searching, feeding, and mating. During rains the grasshoppers have been observed in the Alaskan meadow to take refuge by sitting vertically, head-up on vegetation. Activity, and much resting, may continue until 5 p.m. at which time the grasshoppers begin to bask for a second time. Basking then continues until nearly sunset when they disappear into their nocturnal shelters.
Selected References
Cantrall, I. J. 1943. The ecology of the Orthoptera and Dermaptera of the George Reserve. Michigan Misc. Publ. Mus. Zool. Univ. Michigan, No. 54.
Gurney, A. B. and A. R. Brooks. 1959. Grasshoppers of the mexicanus group, Genus Melanoplus (Orthoptera: Acrididae). Proc. U. S. National Mus. 110: 1-93.
Kreasky, J. B. 1960. Extended diapause in eggs of high-altitude species of grasshoppers, and a note on food-plant preferences of Melanoplus bruneri. Annals Entomol. Soc. Amer. 53: 436-438.
Kaufmann, T. 1971. Biology and ecology of Melanoplus borealis (Orthoptera: Acrididae) in Fairbanks, Alaska with special reference to feeding habits. Michigan Entomologist 4: 3-13.
Vickery, V. R. And D. K. McE. Kevan. 1985. The grasshoppers, crickets, and related insects of Canada and adjacent regions. Research Branch Agric. Canada Publ. 1777.
Larsen, J. C., J. A. Hutchason, T. McNary, and K. Zimmerman. 1988. The Wyoming grasshopper information system. Coop. Agric. Pest Survey, University of Wyoming, Laramie.
Next Species in Subfamily: Melanoplus bowditchi
The University of Wyoming has earned its Research Level 1 (R1) status from the Carnegie Classification of Institutions of Higher Education, placing Wyoming's only four-year university with the top research universities in the United States.