
Grasshoppers of Wyoming and the West
Entomology
Finned Grasshopper
Trachyrhachys aspera Scudder
 |
Distribution and Habitat
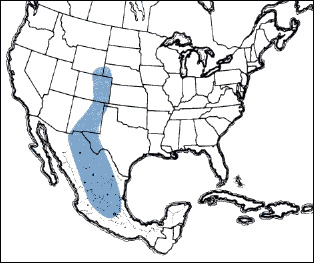 T. aspera continental distribution map >
T. aspera continental distribution map >
The finned grasshopper inhabits the shortgrass prairie and to a limited extent the desert prairie and the western edge of the mixedgrass prairie, ranging from Wyoming and Nebraska to central Mexico. Vegetation of these grasslands consists of the dominant plant, blue grama grass, several subdominant grasses, forbs, small shrubs, and much bare ground. During daylight hours, the finned grasshopper rests and conducts its activities on blue grama plants and on bare ground; at night it rests on bare ground and ground litter under canopies of blue grama.
Economic Importance
Finned grasshoppers feed almost entirely on blue grama grass, the dominant plant in their habitats and a primary forage plant of livestock. In recent years, population densities have been low and no known outbreaks have occurred. Although the Cooperative Economic Insect Survey recorded damaging numbers of the Kiowa grasshopper, Trachyrhachys kiowa, for 19 years between 1951 and 1980, the finned grasshopper was not mentioned once.
Collected from a mixedgrass prairie site in southeast Wyoming, live weight of six males averaged 202 mg and six females 461 mg (dry weight: males 59 mg, females 140 mg). The average dry weight of males and females are similar to those of the bigheaded grasshopper, Aulocara elliotti, a large and injurious rangeland species. Assuming an equal impact by grasshoppers of the same size and a density of one young adult finned grasshopper per square yard, we can estimate an annual loss of 20 pounds (dry weight) of forage per acre. During an outbreak, rangeland sites harbor assemblages of grasshopper species in which the finned grasshopper may be present as a subdominant member. Even the low density of the finned grasshopper during an outbreak contributes to serious damage of forage.
Food Habits
The finned grasshopper feeds almost exclusively on blue grama grass. A thrifty feeder, this grasshopper devours green leaves of blue grama from tip to base. If a leaf is clipped, the grasshopper holds onto the detached section and consumes it entirely. A study of the food habits of this grasshopper in the shortgrass prairie of Colorado revealed that of 13 adults, the crop contents of 12 consisted entirely of blue grama; a single crop had 99 percent blue grama and 1 percent sand dropseed. In the desert prairie of western Texas, crop contents consisted of 80 percent blue grama and 20 percent hairy grama, a closely related species. Two-choice food preference tests of adults collected from a mixedgrass prairie site in southeast Wyoming showed that blue grama was clearly preferred to western wheatgrass, red threeawn, downy brome, and threadleaf sedge.
Observations of feeding behavior of the finned grasshopper were made in a laboratory cage (1 ft3) provided with sod translocated from the mixedgrass prairie of southeast Wyoming. A typical observation was made of a female starved for 19 hours and placed on bare soil in the cage. She crawled around on the bare soil for three minutes before she contacted a blue grama plant. She raised up diagonally on the plant and fed on a green leaf three inches long from tip to base, handling the leaf with her front tarsi while resting on bare soil with mid and hindlegs. She repeated this behavior five times. The sixth leaf she cut at one inch level, held onto it, fed on all of the green portion, and dropped the dry yellow tip. Finally she fed on the stub of blue grama. The time expended searching and feeding on six leaves and one stub was 28 minutes. Another female fed in the same manner, except for picking up an inch section of a fresh blue grama leaf from the soil surface and consuming all of it. She took 18 minutes from start to end of feeding.
Dispersal and Migration
The finned grasshopper's widespread presence in the shortgrass prairie where it inhabits patches of blue grama grass indicates effective dispersal. No doubt the species had many centuries to disperse slowly north upon retreat of the last ice sheet as the climate started to warm some 18,000 years ago. Nevertheless, this grasshopper appears to be a relatively sedentary species that exhibits faithfulness to favored habitats. Flushed flight is short (3 to 4 ft), low (4 to 6 in), and silent. Appetitive flights have never been observed in the two study sites of Colorado and Wyoming. Because these observations lack sufficient replication and completeness, conclusions remain tentative until more is learned about this grasshopper's habits of dispersal.
Identification
The finned grasshopper is a medium size, brown, mottled gray and black grasshopper (Fig. 6 and 7). It often occupies the same habitat as the Kiowa grasshopper which it closely resembles. Several characters serve to separate the two species. The hindwing of the finned grasshopper is marked by a large black band that often extends to the apex (Fig. 9), while the hindwing of Kiowa is mainly transparent. The posteroventral angle of the pronotal lateral lobe of the finned grasshopper approximates a right angle, while that of the Kiowa grasshopper is acute and the lobe distinctly drawn down. The inner (medial) side of the hind femur of the finned grasshopper is chiefly black marked by one light transverse bar (Fig. 8), while the Kiowa grasshopper is marked by two light transverse bars. The hind femur of the finned grasshopper has a high dorsal keel (fin) (Fig. 7), while the Kiowa grasshopper has a low dorsal keel. The finned grasshopper has a thin brush of hairs on the lower keel (Fig. 7), while the Kiowa grasshopper has a dense brush.
The nymphs are identifiable by their shape, structures, and color patterns (Fig. 1-5).
1. Head: face nearly vertical; compound eye with bottom third dark brown and black,
upper two-thirds tan with three dark, diagonal stripes; antennae filiform, proximally
gray, distally black.
2. Pronotum: median carina distinct, incised twice; posteroventral angle of lobe
approximating a right angle.
3. Hind femur with inner face mainly black, one pale yellow bar in front of knee
(Fig. 8); in instar I the outer surface of the hind femur is strikingly black except for
a light area proximally (Fig. 1); hind tibia usually black, sometimes gray.
4. Body short and robust; color brown with black markings, venter generally yellow
or pale tan, and unspotted.
Nymphs of the finned and Kiowa grasshoppers occupying the same habitat present a problem of identification. The easiest way of telling the nymphs apart is to examine the inner face of the hind femur. Nymphs of the finned grasshopper have the inner face chiefly black with one pale bar in front of the knee, while the inner femur of the Kiowa grasshopper is less black and has two pale bars. Also the phenologies of the two species are different; the finned grasshopper hatches and develops about four weeks later in the season, but some of the instars overlap.
Hatching
The finned grasshopper hatches late in the season, approximately four weeks after Kiowa with which it often shares the same habitat and overlaps phenologically. In the shortgrass prairie of north central Colorado (Central Plains Experimental Range, elevation 5,400 ft), hatching usually begins the first week of July and continues for 7 to 10 days. Farther south in Trans-Pecos, Texas, hatching occurs at the same time. This event is delayed in southwest Texas by the sparse amount of spring rainfall and triggered by adequate amounts falling in June and July.
Nymphal Development
In Colorado the nymphs develop during July and the first half of August when temperatures are at their hottest. The host plant of this grasshopper, blue grama, remains green and succulent even though other native grasses may have cured or desiccated. Based on the times of appearance of first instars (hatching) and first adults, the nymphal period lasts 47 to 58 days (average 51 days, 1972 to 1975).
Adults and Reproduction
In northern Colorado the finned grasshopper molts to the adult stage during the last two weeks of August, the males fledging one week ahead of females. At this time, the adult stage is also reached farther south, in Trans-Pecos, Texas (Marathon, Texas elevation 4,121 ft). The adults are present in their habitats from mid August through October.
Oviposition was not observed in nature and rarely in cages, presumably because of the secretive behavior of females. In a cubic foot cage into which blue grama sod had been translocated from the natural habitat, a female was observed drilling and attempting to oviposit for 60 minutes into a small bare area (2 sq in) among grama plants. Search of the soil, however, revealed no eggs. At the end of life of six pairs that had been held individually in six cages in the laboratory, the sod and soil were examined for eggs. Production ranged from 1 to 48 eggs per female and averaged 30. The females laid in the small bare areas between grama plants at depths of 1/4 inch (top eggs of a clutch) to 1 1/4 inches (bottom eggs). No pod was formed, but a very light coating of froth enveloped some of the eggs. Number of eggs in a clutch ranged from 12 to 18. Eggs were 4.6 to 5.2 mm long and yellow to dark brown, some were two-toned yellow and dark brown (Fig. 10).
An interesting result of the cage studies was the lack of oviposition by females in the summer of 1999 when they were exposed daily to 12 hours of light and heat by a 25 watt incandescent bulb and to daylight entering through south-facing windows. In 2000, the 25 watt bulb of each cage was programmed for 7 hours on and 17 hours off with light coming through south-facing windows. Under these conditions each female produced eggs. We venture that like Melanoplus devastator, maturation of T. aspera is triggered by declining day lengths of late summer. Apparently the stimulus of declining photoperiod is also required by Cordillacris crenulata to develop eggs and oviposit, as this species did not produce eggs under 12 hours of daily light during adulthood.
Population Ecology
Collections and observations of the finned grasshopper in Colorado, Texas, and Wyoming indicate that the species is a subdominant member of rangeland grasshopper assemblages and survives perennially at low densities in favorable habitats of shortgrass prairie where blue grama is the dominant grass. In the longest studied site, the Central Plains Experimental Range in north central Colorado, sparse populations inhabited shortgrass prairie annually from 1968 to 1978. From 1972 to 1975 recorded densities ranged from 0.27 to 0.72 young adults per square yard (Table 1), demonstrating that like other species of grasshoppers, fluctuations in population density occur. Subsequent research detected populations from 1980 to 1986 and in 1989 and 1999. In intervening years this rangeland was not searched for grasshoppers. The evidence indicates a permanent occupation of this shortgrass prairie site by sparse populations of the finned grasshopper.
Distribution of the finned grasshopper is spotty in the north of its range, with populations inhabiting limited acreage of shortgrass prairie. In the study sites in southeast Laramie County, Wyoming, the population was localized in an apparently favorable habitat of several acres. In contrast to other species of grasshoppers, such as Trachyrhachys kiowa which inhabited a much larger area outside this favorable site, T. aspera was notably absent. A New Mexico dot map indicates that near the center of its geographical range this grasshopper is widely distributed over shortgrass prairie, however even here densities have remained low.
Calculated daily mortality rates of adults were low (approximately 2 percent) allowing this stage to survive into late fall—through September and October. Why the species remains numerically low and a subdominant member of the grasshopper assemblage is an interesting and important question. One possibility is that females have a low fecundity in nature like in our cage study. Another possibility is that high egg mortality may occur during the period following deposition, a time of minimal rainfall. Because the eggs lie loosely in the soil unprotected by a pod, they may be at great risk of desiccating.
Daily Activity
At night finned grasshoppers take shelter on the ground surface under canopies of blue grama grass. In late August they remain hidden until two hours after sunrise at which time they crawl out onto bare ground and begin to bask turning a side perpendicular to the rays of the sun and lowering the hindleg to expose the abdomen. They bask for 90 minutes to two hours. During this time, approximately 8 to 10 am DST, the temperatures of the ground surface may rise from 50°F to 115°F. Still on the bare soil, they continue sitting, but take postures that reduce insolation and may even begin to stilt. In the laboratory study, males during late morning were observed crawling about on blue grama while females usually remained quiet and hidden from view in the thick of the grass. At various times during the day, caged males were observed mounted on females and in copula. Courtship was brief; stridulating males approached females and when close, approximately 1/2 inch away, they jumped onto the back of females and attempted to mate. When receptive females were mounted, mating followed. In the southeast Wyoming study site, a male was observed having mounted a female on 21 August 2000 at 11:27 am. A caged pair was observed in copula on 26 August 2000 at 11:30 am.
Finned grasshoppers appear to have low heat tolerance. On clear days they begin to stilt at 10:40 am (ground temperature 115°F, air 73°F). Soon afterwards they climb on top of grama grass, approximately 1 inch above the soil surface; then face the sun directly so that only the front of the head is impinged while the rest of the body is shaded. They were observed to avoid high temperatures from morning to late afternoon (10:40 am to 4:30 pm). During this time no other voluntary activities were observed. The question arises: when do these grasshoppers feed? A female translocated onto rangeland grama grass fed after sunset (6:56 pm, 14 September 2000, soil temperature 68°F, air temperature 67°F). Evidently finned grasshoppers can make adjustments in the time of their feeding to the temperature flux of their habitat. Intensive investigation will be required to discover all the details of their daily activities.
Selected References
Capinera, J.L. and D.C. Thompson. 1987. Dynamics and structure of grasshopper assemblages in shortgrass prairie. Can. Entomol. 119: 567-575.
Joem, A. 1979. Resource utilization and community structure in assemblages of arid grassland grasshoppers (Orthoptera: Acrididae). Trans. Amer. Entomol. Soc. 105: 253-300.
Joern, A. 1982. Vegetation structure and microhabitat selection in grasshoppers (Orthoptera: Acrididae). Southwestern Naturalist 27: 197-209.
Pfadt, R.E. and R. L. Lavigne. 1982. Food habits of grasshoppers inhabiting the Pawnee site. Wyoming Agr. Exp. Stn. SM 42.
Richman, D.B., D.C. Lightfoot, C.A. Sutherland, and D.J. Ferguson. 1993. A manual of the grasshoppers of New Mexico, Orthoptera: Acrididae and Romaleidae. New Mexico State University Handbook 7.
Next Species in Subfamily: Trachyrhachys kiowa
The University of Wyoming has earned its Research Level 1 (R1) status from the Carnegie Classification of Institutions of Higher Education, placing Wyoming's only four-year university with the top research universities in the United States.